| Operon laktozowy | Promotory | Transkrypcja | Strona g��wna | Bia�ka i ich funkcje | Kontakt |
| Operon laktozowy |
| Promotory |
| Transkrypcja |
| Strona g��wna |
| Bia�ka i ich funkcje |
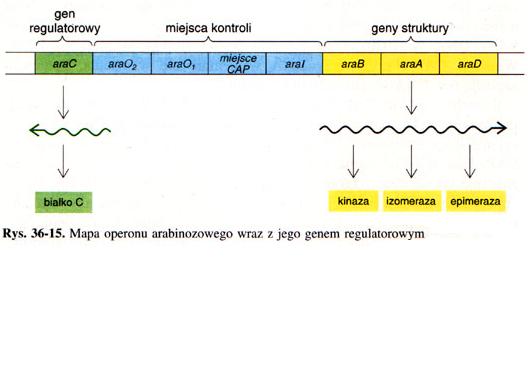
Operon arabinozowy
R�ne formy tego samego bia�ka aktywuj� lub blokuj� transkrypcj� operonu
arabinozowego.
Bakteria mo�e korzysta� z arabinozy jako �r�d�a energii przekszta�caj�c j� w
ksylulozo-5-fosforan, kt�ry z kolei ulega przekszta�ceniu w intermediaty
glikolizy. Ksylulozo-5-fosforan powstaje z arabinozy w wyniku kolejnego
dzia�ania izomerazy arabinozowej, rybulokinazy i
epimerazy rybulozo-5-fosforanowej. Enzymy te s� kodowane odpowiednio
przez geny araA, araB, araD, nale��ce do
operonu arabinozowego. Operon ten r�wnie� zawiera gen bia�ka regulatorowego
C (araC), dwa miejsca operatowe
(araO1, araO2), miejsce wi�zania dla CAP oraz
inne miejsca regulatorowe araI.
Operon arabinozowy, tak jak inne indukowane operony kataboliczne, podlega
podw�jnej kontroli. Do wydajnej transkrypcji konieczne s� dwa sygna�y:
kompleks cAMP z CAP, oraz arabinoza zwi�zana z bia�kiem C. To ostatnie jest
r�wnie� negatywnym regulatorem genu araC. Transkrypcja araC, kt�ra zachodzi
w kierunku przeciwnym do transkrypcji araBAD, jest kontrolowana przez
miejsce operatorowe, O1, a nie przez O2. Synteza mRNA bia�ka C zachodzi gdy
st�enie tego bia�ka i kompleksu cAMP-CAP jest ma�e. Kiedy natomiast jest
du�o bia�ka C i nie ma cAMP-CAP, transkrypcja genu bia�ka C jest zahamowana,
poniewa� bia�ko C wi��e si� z araO1. synteza bia�ka C podlega autoregulacji.
Przy��czenie drugiej cz�steczki bia�ka C do araO2 blokuj� syntez� mRNA BAD
przez utworzenie p�tli DNA i zwi�zanie si� z araI, kt�re to miejsce
s�siaduje z promotorem gen�w araBAD. Wyp�tlanie zapobiega przy��czeniu
polimerazy RNA.
P�tla DNA nie tworzy si� kiedy jest du�o kompleksu
cAMP-CAP i arabinoza wi��e si� z bia�kiem C
zmieniaj�c jego konformacj�. Obecno�� tych pozytywnych czynnik�w
regulatorowych umo�liwia polimerazie RNA przy��czenie si� do promotora gen�w
BAD i ich transkrypcj�. W tej sytuacji mRNA bia�ka C nie powstaje poniewa�
araO1 jest zaj�te przez bia�ko C.
Operon arabinozowy ilustruje kilka zasadniczych cech regulacji gen�w.
Po pierwsze bia�ko mo�e samo regulowa� w�asn�
syntez� przez represj� transkrypcji w�asnego genu. Po
drugie wi�zanie cz�steczki sygna�owej do bia�ka mo�e przekszta�ci�
bia�ko w inhibitora transkrypcji w aktywator. Odmienne formy tego samego
bia�ka wi��� si� do r�nych miejsc regulatorowych w DNA.
Po trzecie miejsca wi���ce bia�ka regulatorowe
w DNA nie musz� s�siadowa� z kontrolowanymi przez nie genami. Operon
arabinozowy dostarcza przyk�ady modulowania transkrypcji w miejscach
znajduj�cych si� w pewnej odleg�o�ci od transkrybowanego genu. DNA mo�e by�
"zap�tlony" przez zwi�zanie bia�ka lub kompleksu bia�kowego z miejscami DNA,
kt�re ze sob� nie s�siaduj�. Wyp�tlanie DNA cz�sto wyst�puj� w regulacji
gen�w, szczeg�lnie u eukariot�w. Po czwarte
zmiany indukowane przez cz�steczk� sygna�ow� s� �atwo odwracalne. System
odpowiada ci�gle i szybko na zmiany st�enia metabolit�w.